För att fåglar överhuvudtaget skall kunna överleva längre perioder i havsmiljö måste de kunna göra sig av med det överskottssalt, som de får i sig då de dricker och äter. Utsöndringen av överskottssaltet sker via en s.k. saltkörtel belägen vid näbbens ovansida. Dan-Axel Hallbäck beskriver här saltkörtelns uppbyggnad och funktion.
Pelagiska fåglar, d.v.s. fåglar som längre tid vistas över öppna havsvidder utan att ha kontakt med land, har framför allt karakteriserat jordklotets södra oceaner. Under de senaste hundra åren har dock Nordatlantens vatten berikats med en art, stormfågeln Fulmarus glacialis, som nästan explosionsartat expanderat och fyllt en ledig ekologisk nisch. Före 1878 fann man den endast i rent arktiska områden; förutom två mycket små kolonier på Island respektive St. Kilda väster om Hebriderna (Fisher 1952). En av förutsättningarna för artens expansion, liksom för att fåglar överhuvudtaget skall kunna leva på öppna havsvidder utan tillgång till sötvatten, är förmågan att ta upp saltvatten (salthalt 35-50 %o), behålla vattnet och göra sig av med överskottssaltet. Kunskapen om hur detta går till är av relativt ungt datum, och det första moderna arbetet kom så sent som 1957. Skandinaviska forskare som Knud Schmidt-Nielsen och Ragnar Fänge har i hög grad bidragit till förståelsen av dessa för havsfåglar så vitala funktioner.
Man visste redan tidigare, att fåglars njurar är dåligt utvecklade vad gäller förmågan att koncentrera urin. I detta avseende liknar fågelnjuren mer kräldjurens än den synnerligen effektiva daggdjursnjuren, som hos framför allt ökenlevande djur drivits mycket långt i frågan om fysiologisk och ekologisk anpassning och effektivitet (Dicker 1970). Hos marina fåglar kom därför intresset tidigt att knytas till en liten körtel vid näbbens ovansida. Rent anatomiskt beskrevs den för första gången (hos tamand) redan på ett anatomimöte i Amsterdam den 14 november 1665 (Peaker & Linzell 1975). Denna beskrivning har tillskrivits en forskare vid namn Caspar Commelin, men vem han egentligen var är okänt (Anon. 1667). Mer i detalj beskrevs körteln av Jakobson (1813) och av Nitzsch (1820). Uppbyggnad och funktion. Saltkörteln kan vara belägen framför, i eller runt ögonhålan. Hos utpräglat marina fåglar är den stor och ligger i en speciell benficka (Technau 1936).

The arrangement of lobes in a salt gland of Herring Gull (from Fänge et al. 1958).
Körteln kan variera i storlek och består av 4-25 lober. Generellt kan sägas, att ju mer marin en fågelart är, desto större är körteln och ju fler lober är den uppbyggd av. Varje lob har en centralt belägen gång, och dessa gångar samlas till en eller flera utförsgångar, som mynnar i näshålan (figur 1). Saltkörteln får sin blodförsörjning från de inre och yttre synartärerna samt från näbbartären (figur 2). De tre kärlen har förbindelser sinsemellan. Som exempel kan nämnas, att hos gråtrut Larus argentatus är dessa artärer bland de grövsta i fågelns huvud, vilket anses bero på att blodflödet tillen saltkörtel under maximal utsöndring är mycket stort (Fänge m.fl. 1958). Jämför man blodflödena till saltkörteln och hjärtmuskeln

- Figur 2. Den arteriella försörjningen av saltkörteln hos gråtrut (från Fänge m.fl. 1958).
The provision of arterial blood to the salt gland of Herring Gull (from Fänge et al. 1958).
visar det sig, att flödet till en körtel under maximal utsöndring är ungefär tre gånger så stort som till hjärtat, räknat per 100 g organvikt (Hanwell m.fl. 1971a, 1971b). Venerna från saltkörteln följer i allmänhet artärerna. Vad gäller körtelns lymfdränage är fortfarande mycket litet känt (Peaker & Linzell 1975).
Saltkörteln innerveras från flera nerver, och de viktigaste är: Nerver från V:e hjärnnerven N. trigeminus, en speciell nerv benämnd ”the secretory nerve” (innehåller flera fibrer från VII:e och IX:e hjärnnerverna, N. fascialis resp. N. glossopharyngeus), samt nerver från det sympatiska nervsystemet.
Med ovanstående grova anatomiska skiss som bakgrund skall vi nu gå vidare och försöka göra oss en bild av hur saltkörteln fungerar. Först måste vi då titta lite närmare på hur en lob är uppbyggd (figurerna 1 och 4). Som tidigare sagts, löper en central kanal mitt i loben. I denna kanal tömmer sig ett stort antal utsöndringsrör. Rören är radiärt anordnade runt centralkanalen och slutar blint i sin yttre ända. Utsöndringsrören är klädda med speciella celler (figur 3), som mycket liknar cellerna i andra utsöndrande ytorgan.

Detail of the micro circulation in a salt gland lobe (from Fänge et al. 1958).
Cellerna kännetecknas dels av en kraftig veckbildning basalt, med en kraftig förstoring av ytan mot kroppvätskorna som följd, dels av en stor rikedom på mitokondrier (mitokondrierna är cellorganeller som framförallt svarar för energiproduktionen). Den rika förekomsten av mitokondrier förklaras av att transporten av saltjoner från kroppsvätskorna till de utsöndrande rörens hålig-heter är en extremt energikrävande process. Om man låter marina fåglar få fri tillgång till sötvatten finner man, att de banala vecken slätas ut och att antalet mitokondrier minskar, eftersom saltkörteln, så att säga, går på sparlåga (Ernst & Ellis 1969). Utsöndringscellerna är således basen för utsöndringen av salt, men det finns andra mekanismer, som hjälper till med att producera den koncentrerade saltlösning, som blir resultatet av körtelns verksamhet.

The ultra structure of an excretory cell in an active salt gland (from Peaker & Linzell 1975 with permission from the authors and Cambridge Univ. Press).
En sådan mekanism visas i figur 4: En eller två små artärer löper parallellt med det utsöndrande röret, men flödesriktningarna i artärer respektive rör är motsatta. Denna och andra detaljer i körtelns uppbyggnad innebär förutsättningar för ett utbyte av ämnen mellan de bägge flödena (s.k. motströmsutbyte). Det arteriella blodet, som kan antas ha ett högt saltinnehåll sedan fågeln druckit saltvatten, når utsöndringsrören i en punkt, där innehållet i rörens håligheter är som störst. Ju mer perifert man når, desto saltfattigare blir innehållet i såväl blodet som i rörlumen. Med andra ord, koncentrationsskillnaden mellan artär och utsöndringsrör är (teoretiskt) konstant, och detta innebär att hela rörlängden kan utnyttjas för salttransport. Man har också funnit, att längden av utsöndringsrören är direkt proportionell till saltkörtelns förmåga att koncentrera sekretet(Staaland 1967, 1968). Effektiviteten i utsöndringsarbetet ökas på detta sätt i hög grad. Saltkörtelns motströmsutbyte har diskuterats bland annat i en artikel av Schmidt-Nielsen (1960).
Nervös och hormonell styrning av saltkörtelns arbete
Hur kan då saltkörteln ”veta”, när fågeln behöver göra sig av med salt? För detta krävs information, vilken fågeln får via sinnesceller som reagerar på förändringar i kroppsvätskornas volym eller salthalt. Stewart (1972) presenterar försök, där han visar, att det är framförallt ökad salthalt, som startar aktiviteten i saltkörteln hos marina fåglar (trutar). Informationen från sinnescellerna, som i första hand är lokaliserade till hjärtat, förs vidare via nerver till det centrala nervsystemet. Där omkopplas impulserna och integreras med annan information, varefter informationen går vidare via nervknutar ut till cellerna i saltkörtelns utsöndringsrör (figur 5).

A sketch of the nervous regulation of the salt gland (from Hanwell et al. 1972).
Det har ofta hävdats, att styrning via nerver svarar för snabba förändringar i fysiologiska funktioner (beträffande t.ex. blodflöde, körtelfunktioner och hjärtaktivitet), medan mer långsiktiga inställningar framförallt sker med hjälp av hormoner. Ett stort intresse har därför varit knuten till olika hormoners effekter på saltkörteln. Bland de hormon, som studerats mest i detta avseende, kan nämnas binjurebarkshormoner (ACTH från hypofysen och olika varianter av kortison från binjuren). Dessa hormoner har visat sig ha en viss effekt på saltkörtelns sekretion, men det troligaste är att detta är bieffekter till den ofta dramatiska påverkan, som dessa hormon har på hela kroppens inre kemiska miljö.
Hypofysen är ett annat hormonproducerande organ av stort intresse. Den består av två delar, en främre och en bakre lob, den senare även kallad neurohypofysloben. Bland framlobens hormon har speciellt prolaktin (som deltar i styrningen av mjölkproduktion hos människor) studerats. Undersökningar visar, att prolaktin kan påverka saltkörtelns funktion så snabbt efter hormonstimulering, att effekterna rimligtvis inte kan förklaras som allmänna omställningar. Huruvida prolaktin verkar direkt på saltkörteln eller via andra mekanismer är dock oklart. För tydlighetens skull bör man kanske tillägga, att prolaktin är ett utvecklingsmässigt sett gammalt hormon, vilket t.ex. hos en del sötvattensfiskar fungerar genom att stimulera jontransport över cellmembran.
Baklobens (neurohypofysens) ”klassiska” hormon, såsom vattensparande hormon (ADH), tillväxtstimulerande hormon (STH) samt hormonet MSH (reglerar hudens innehåll av pigment), har inte något större inflytande på saltkörteln. Däremot finns ett speciellt hormon, AVT (arginine vasotocin), som eventuellt kan spela en roll för långtidsstyrningen av saltkörteln.
Sammanfattningsvis kan man alltså säga, att visserligen har många hormon en positiv eller negativ effekt på saltkörtelns arbete, men någon direkt styrande effekt av hormoniell karaktär har man ännu inte kunnat påvisa (för referenser beträffande hormoniell styrning, se Peaker 1971).
Utveckling och förekomst hos olika djurgrupper
Saltkörtlar finner man enbart hos högstående reptiler och hos fåglar, och från fossilfynd kan man sluta sig till, att de kan ha uppstått ur amfibiernas s.k. nasalkörtlar. Bildandet av saltkörtlar har satts i samband med att två viktiga jonreglerande organ, gälarna och den tunna amfibiehuden, gick förlorade när kräldjuren uppstod. Härtill kommer, att när djuren började ta landbacken i besittning tvingades de övergå från en växtdiet med ungefär samma jonkoncentration som havet till landlevande växter med en annan jonuppsättning. Troligen var de tidiga landväxterna bl.a. mer kaliumrika. Det är således möjligt, om man skall tro Sokol (1967), att de första saltkörtlarna utvecklades för att reglera kroppens natrium-kaliumbalans och inte i första hand för att spara vatten.
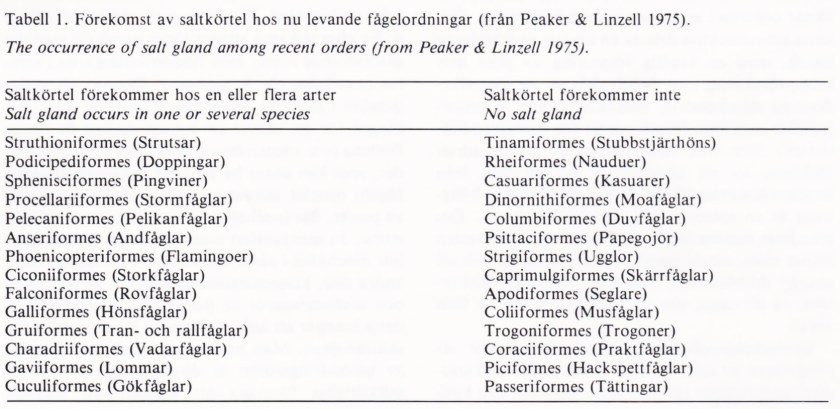
Saltkörtelns funktion hos dagens fåglar varierar från släkte till släkte och har anpassats till skilda ekologiska krav. Förekomst av saltkörtel har konstaterats hos ungefär hälften av de nu levande fågelordningarna (tabell 1).
Hos ökenlevande fåglar är saltkörteln stor och välutvecklad. Speciellt gäller detta unga individer, medan funktion och betydelse tycks avta med åldern. Förklaringen till detta är delvis ekologisk. Vuxna ökenfåglar lever som regel ej helt utan tillgång på vatten, även om de kan tvingas att flyga ansenliga sträckor till närmaste vattenhål. Så länge ungarna är kvar i boet eller dess närhet är de emellertid hänvisade till den vätska, som de kan få på platsen, framförallt via sina föräldrar. Dessutom är ungarna ofta utsatta för en brännande solhetta med stora förluster av vatten som följd. Då saltkörteln producerar en vätska med högre saltinnehåll än fågelns blodplasma, kan fågelns göra sig av med överskottssaltet och balansera avdunstningen av vatten (Ohmart 1972).

De vackraste exemplen på saltkörtlar finner man emellertid hos havslevande fåglar; framförallt hos albatrosser, stormfåglar, liror och stormsvalor, men också hos alkor, måsfåglar samt vissa änder och gäss. Hos dessa fåglar är körtelns enda funktion att utsöndra överskottet av koksalt. Marina fåglar kan på grund av denna körtel dricka saltvatten utan att riskera saltförgiftning. På så sätt har förutsättningar skapats för ett pelagiskt liv, som tillåter fåglarna att leva halvårsvis ute till havs och därmed tillgodogöra sig resurser i den ”ekologiska nisch”, som rent rumsligt täcker 70 Wo av jordklotets yta.
Summary: The salt gland – an essential organ of marine birds
Bird species of different orders have evolved a nasal gland for the excretion of surplus salt (Tab. 1), although this salt gland occurs most frequently among marine and desert living species. The gland consists of 4-25 lo-bes, each with a central excretion channel (Fig. 1), and is supplied with blood from three large arteries, allowing a large blood flow (Fig. 2). A large number of excretion tubes drain into the central channel of each lobe; the walls of these tubes are covered with excretory cells (Fig. 3). These cells are supplemented with a counter-current system for the exchange of salt ions between the blood and the liquid in the excretion tubes (Fig. 4). The function of the salt glands is regulated by nerves (Fig. 5), while hormonal control seems to be of minor importance.
Tabell 1. Förekomst av saltkörtel hos nu levande fågelordningar (från Peaker & Linzell 1975). The occurrence of salt gland among recent orders (from Peaker & Linzell 1975).
Saltkörtel förekommer hos en eller flera arter Saltkörtel förekommer inte
Salt gland occurs in one or several species No salt gland
Struthioniformes (Strutsar) Podicipediformes (Doppingar) Sphenisciformes (Pingviner) Procellariiformes (Stormfåglar) Pelecaniformes (Pelikanfåglar) Anseriformes (Andfåglar) Phoenicopteriformes (Flamingoer) Ciconiiformes (Storkfåglar) Falconiformes (Rovfåglar) Galliformes (Hönsfåglar) Gruiformes (Tran- och rallfåglar) Charadriiformes (Vadarfåglar) Gaviiformes (Lommar) Cuculiformes (Gökfåglar)
Tinamiformes (Stubbstjärthöns) Rheiformes (Nauduer) Casuariformes (Kasuarer) Dinornithiformes (Moafåglar) Columbiformes (Duvfåglar) Psittaciformes (Papegojor) Strigiformes (Ugglor) Caprimulgiformes (Skärrfåglar) Apodiformes (Seglare) Coliiformes (Musfåglar) Trogoniformes (Trogoner) Coraciiformes (Praktfåglar) Piciformes (Hackspettfåglar) Passeriformes (Tättingar)
Litteratur
Anon. 1667. Observationes anatomicae selectiores collegi privati Amstelodamensis. Apud C. Commelinum. Amstelodami. (Referensen finns i Cole (1938) och Peaker & Linzell (1975)).
Cole, F.J. 1938. Observtiones anatomicae selectiores Amstelodamensium 1667-1673. University of Reading, Reading.
Ernst, S.A. & Ellis, R.A. 1969. The development of surface specialization in the secretory epithelium of the avian salt gland in response to osmotic stress. Journal of Cell Biology 40: 305-321.
Fisher, J. 1952. The Fulmar. The New Naturlaist, monograph No. 6. Collins, London.
Fänge, R., Schmidt-Nielsen, K. & Osaki, H. 1958. The salt gland of the herring gull. Biological Bulletin, Marine Biological Laboratory, Woods Hole, Mass. 115: 162-171.
Hanwell, A., Linzell, J.L. & Peaker, M. 1971a. Salt gland secretion and blood flow in the goose. Journal of Physiology 213: 373-387.
Hanwell, A., Linzell, J.L. & Peaker, M. 1971b. Cardiovascular response to salt-loading in conscious domestic geese. Journal of Physiology 213: 389-398.
Hanwell, A., Linzell, J.L. & Peaker, M. 1972. Nature and location of the resoptors for salt secretion in the goose. Journal of Physiology 226: 453-472.
Jacobson, L.L. 1813. Sur une glande conglomée appartenante å la cavité nasale. Bulletin de la Soci Philomathique de Paris 3: 267-269.
Nitzsch, C.L. 1820. Uber die Nasendruse der Vogel. Deutsches Archiv fur die Physiologie (Meckels Archiv) 6: 234-269.
Ohmart, R.D. 1972. Physiological and ecological observations concerning the salt-secreting nasal glands of the roadrunner. Comparative Biochemistry and Physiology 43 A: 311-316.
Peaker, M. 1971. Avian salt glands. Philosophical Transactions of The Royal Society of London B 262: 289-300.
Peaker, M. & Linzell, J.L. 1975. Salt glands in birds and reptils. Monographs of The Physiological Society No. 32. Cambridge University Press, Cambridge.
Schmidt-Nielsen, K. 1960. The salt-secreting gland of marine birds. Circulation 21: 955-967.
Sokol, O.M. 1967. Herbivory in lizards. Evolution 21: 192-194.
Staaland, H. 1967. Anatomical and physiological adaptations of the nasal glands in Charadriiformes birds. Comp. Bioche. Physiol. 23: 933-944.
Staaland, H. 1968. Excretion of salt in waders, Charadrii, after acute salt loads. Nytt Magasin for Zoologi 16: 25-28.
Stewart, D.J. 1972. Secretion by salt gland during water deprivation in the duck. American Journal of Physiology 223: 384-386.
Technau, G. 1936. Die Nasendruse der Vogel. Journal fur Ornithologie 84: 511-617.
Publicerad i Vår fågelvärld nr 42 (1983)
